
Rev. Fac. Agron. (LUZ). 1997, 14: 193-206
Biología de la mosca blanca del tabaco, Bemisia tabaci (Homoptera: Aleyrodidae) y potencial para desarrollar sus poblaciones sobre cinco especies de plantas hospederas.1
Bionomics of the tobacco whitefly, Bemisia tabaci (Homoptera: Aleyrodidae) and potential for population increase on five host plant species.
l Aceptado
el 04-07-1996
1. Trabajo realizado con financiamiento de la Fundación para el Desarrollo de la Ciencia
y la Tecnología (FUNDACITE-Zulia).
2. Departamento de Botánica. Facultad de Agronomía. L.U.Z.
3. Unidad Técnica Fitosanitaria (U.T.F.) Facultad de Agronomía. La Universidad del
Zulia. Maracaibo, ZU 4005, Venezuela.
4. Departamento de Estadística. Facultad de Agronomía. L.U.Z.
Adriana Sánchez2 , Francis Geraud-Pouey3 y
Douglas Esparza4
Resumen
Palabras claves: Bemisia tabaci, parámetros demográficos, simulaciones
poblacionales.
Abstract
Key words: Bemisia tabaci, demographic parameters, population simulations.
Introducción
La mosca blanca del tabaco (MBT), Bemisia tabaci (Gennadius), Homoptera:Aleyrodidae, ha sido conocida en Venezuela por muchos años (19). Fuera de su asociación con la transmisión de enfermedades virales de plantas (13, 23, 30), no era considerada de importancia como plaga, por sus usualmente bajas poblaciones. B. tabaci es una especie fitofaga, distribuida ampliamente en las zonas tropicales y subtropicales en todos los continentes (21, 25) y ha sido reportada sobre más de 500 especies de plantas hospederas (18). En Venezuela se le conocen 27 hospederas (2).
A partir de 1988, se presentaron serios problemas debido a altas poblaciones y consecuentes daños en varios cultivos de diferentes regiones del país (16). Cambios de importancia de esta especie como plaga, también ocurrieron en otras regiones del mundo (7, 8, 12), a veces asociado con intenso uso de insecticidas (15). Sin embargo, la existencia de una nueva especie muy cercana, B. argentifolia (4), deja confusa la responsabilidad de cada una de ellas en este problema entomológico. Para precisar la situación taxonómica de la MBT en Venezuela, actualmente se está realizando una evaluación genética de especímenes recolectados en el país (Mario Cermeli, CENIAP-FONAIAP, Maracay, estado Aragua, Venezuela; comunicación personal).
A pesar de su reconocida polifagía, en el campo se han observado diferencias de infestaciones y daños dependiendo de la especie de planta hospedera. En lo referente a la relación insecto-planta, este efecto puede tener dos componentes; en primer lugar, cuan adecuada es la planta para la supervencia, desarrollo y reproducción del insecto fitófago, lo cual en gran parte define el potencial para aumentar sus poblaciones. En el caso de Bemisia esas respuestas pueden estar relacionadas con la especie de planta (12, 31), llegando a variar dentro de la planta de acuerdo a la especie y/o cultivar (31), así como por efecto de edad, posición y superficie de las hojas (7, 24, 28), de manejos agronómicos tales como fertilización (tipo y dosis de fertilizante) (5) o residuos de plaguicidas (24).
Dada la importancia agrícola de este insecto, en lo que atañe a la relación artrópodo fitógago-planta hospedera, es conveniente estudiar su biología sobre algunas de las especies de hospederas cultivadas, para conocer el papel que juega la planta en los problemas entomológicos causados por B. tabaci, en nuestras condiciones. Un segundo componente de este problema, es el efecto que la planta pueda tener sobre la interacción del fitófago y sus enemigos naturales (interacción de tres niveles tróficos) (26). Ello, sería tópico de posteriores estudios.
Materiales y métodos
La investigación, se realizó durante el período julio 1992julio 1993, bajo condiciones de laboratorio [27.1 ± 1.3 °C y 75.5 ± 10.6 % HR, iluminado artificialmente combinando luz fluorescente (12 tubos de 40 watts) e incandescente (6 bulbos de 60 watts) sobre un área de 20 m2, durante aproximadamente 11 h/día, sin excluir la luz natural] en la Unidad Técnica Fitosanitaria (U.T.F.), Facultad de Agronomía, La Universidad del Zulia, Maracaibo, Venezuela.
Las plantas hospederas utilizadas fueron: caraota negra (Phaseolus vulgaris L.), tomate (Lycopersicon esculentum Mill., cv. Rio Grande), algodón (Gossypium hirsutum L., tipo Deltapine), cayena (Hibiscus rosasinensis L., flores rojas simples y hojas lisas) y la planta de navidad o «poinsettia» (Euphorbia pulcherrima L.), las cuales fueron propagadas a partir de semillas o estacas, según el caso, utilizando macetas plásticas con capacidad aproximada para 1 kg de suelo (2 partes de arena limosa + 1 parte de materia orgánica vegetal descompuesta). A los primeros quince días de su desarrollo, se les añadió una fórmula completa de fertilizante (151515; 1 gr/planta), seguido de urea (igual dosis), con una frecuencia quincenal. Las plantas fueron mantenidas al aire libre, agrupadas dentro de jaulasumbráculos estructurada con viguetas (perfiles) de aluminio cubiertas con malla del mismo material (8 hilos/cm) de dimensiones de 2.3 x 1.1 x 1 m (largo x ancho x alto), cuyas paredes largas estaban divididas en dos secciones corredizas; colocadas sobre mesones de 0.9 m de altura.
Los adultos de MBT utilizados para iniciar los experimentos fueron obtenidos de una colonia establecida en el laboratorio, sobre plantas de navidad mantenidas dentro de jaulas entomológicas de madera, con manga de tela (diámetro = 15 cm) en el centro de la puerta frontal, de 32 x 37 x 39 cm (largo x ancho x alto), tope de vidrio en bisel y fondo aireado recubierto con organdí sintético (35 x 27 cm).
Al momento de iniciar los experimentos, las plantas tenian 10-12 hojas verdaderas (40-60 cm de altura). Después de minuciosamente revisadas para eliminar cualquier artrópodo sobre ellas, las plantas fueron colocadas en el piso del cuarto de cría, sin aislamiento individual, solo se le untó vaselina alrededor de las macetas para impedir el acceso a artrópodos rastreros. Para cada una de las especies de plantas hospederas, fueron consideradas tres edades de hojas (jóvenes, maduras y senescentes), 2 hojas/edad/planta. La edad de la hoja fue estimada por su posición en la planta de acuerdo al patrón de crecimiento de cada especie. Sobre cada hoja se colocaron tres hembras y un macho de MBT, confinados dentro de pequeñas jaulas plásticas (microjaulas). La microjaula consistió de un envase cilíndrico (5x3.5 cm, altura x diámetro) de plástico semitransparente, cuya abertura estaba incrustada en una base cuadrada (7 x 7 cm) de cartón corrugado (0.7 cm de espesor), a través de un agujero de igual diámetro, perforado en el centro. La cara inferior de la base (en contacto con la hoja), estaba recubierta por «goma espuma» (esponja sintética, 0.5 cm de espesor). Este componente era colocado sobre el envés de la hoja oponiéndosele por el haz otra lámina de cartón corrugado con «goma espuma». Ambas piezas eran sostenidas mediante 2 pinzas con resorte para cabellos (4 cm de longitud) y ese conjunto soportado mediante una varilla de alambre galvanizado grueso, incrustado en el suelo dentro de la maceta. Los adultos de MBT, eran introducidos a la microjaula, a través de un agujero, (1.2 cm de diámetro) perforado en el fondo del envase, el cual era seguidamente taponeado con algodón. El área foliar expuesta fue de 7 cm2 y el tiempo de exposición de 24 h.
Una vez retiradas las microjaulas con los adultos, se contó el número de huevos. Cada grupo de huevos/microjaula constituyó una unidad experimental y el total de huevos puestos en una fecha, representó una cohorte (grupo de individuos de edad uniforme). Fueron evaluadas seis cohortes incluyendo una planta de cada especie. Mediante observaciones diarias, a cada huevo se le siguió su evolución hasta llegar a adulto, registrándose supervivencia, duración de las fases de desarrollo, sexo y longevidad del adulto emergido. Aquellas hojas que se secaron antes de que los insectos ubicados sobre ellas completaran su desarrollo, no fueron consideradas para los cálculos.
Completado el desarrollo ninfal, las ninfas de cuarto instar (N4) en su fase final («pupas»; cuerpo subgloboso con el dorso de color blanquesino) fueron cuidadosamente despegadas de las hojas usando un alfiler entomológico Nº 1 y mediante un pincel fino colocadas individualmente dentro de cápsulas Petri de plástico transparente (5x1 cm, diámetro x alto) hasta la emergencia de los adultos. Después de sexados los adultos, fueron confinadas parejas (de ambos sexos) dentro de envases plásticos (11 x 13 cm, diámetro x alto; una pareja/envase), conteniendo una hoja de la especie de planta y edad correspondiente a aquella sobre la cual se desarrollaron. Para mantener turgente a la hoja, su pecíolo se mantuvo sumergido en agua dentro de un tubo plástico tipo «vial» (1.5 x 4.5 cm, diámetro x alto), taponeándolo con algodón alrededor del peciolo. Diariamente, fueron cambiadas las hojas y se contó el número de huevos puestos. De esta manera se determinó la fertilidad (número de huevos/hembra/día) durante su vida reproductiva así como la longevidad de los adultos. El diseño experimental utilizado fue un completamente al azar, con un arreglo factorial 5 x 3 (especies de plantas x edades de hoja) y seis repeticiones en el tiempo (cohortes).
Mediante analisis de tablas de vida y fertilidad para cohortes, fueron calculados los parámetros demográficos: tiempo generacional (T = período promedio transcurrido entre el inicio de la generación parental y la generación filial); tasa neta reproductiva (Ro = tasa de multiplicación por generación; individuos por individuo) y tasa intrínseca de desarrollo poblacional (rm= contribución diaria de cada individuo al desarrollo poblacional; individuos por individuo por día) (1, 22). Para ello se utilizó un programa computarizado el cual toma en cuenta el desarrollo variable entre individuos y sexos, combinando estos factores en un cálculo más real de rm (9).
Para detectar el efecto de especie de planta, edad de las hojas y de la interacción de estos dos factores, se utilizó el procedimiento GLM y se hicieron comparaciones de medias de las variables estudiadas, utilizando el método de Tukey para los efectos principales (27), así como la distribución t de Student (29), para las duraciones de los diferentes estadíos, discriminados por sexo.
Con los valores de rm, se realizaron simulaciones del potencial de desarrollo de población de la MBT en el tiempo, sobre cada especie de planta hospedera, utilizando la fórmula: Nt = No ´ erm ´ t (1, 11); donde: Nt: número de individuos al tiempo t, No: número de individuos al inicio de la población (1 individuo/hoja), e: base de los logarítmos neperianos, rm: tasa intrínseca de desarrollo poblacional y t: tiempo para el desarrollo de la población (90 días). Las simulaciones fueron realizadas utilizando el programa Lotus 123Ò (Lotus Develpment Corp.).
Resultados y discusión
Duración del desarrollo. El promedio general para la duración de los huevos fue de 6.2 ± 1.9 días. La duración varió ligeramente sin significancia estadística sobre las diferentes plantas hospederas, siendo el orden tomate (6.5 ± 0.8), caraota (6.2 ± 1.7), cayena (6.2 ± 2.1), navidad (6.0 ± 1.6) y algodón (6.0 ± 2.2). Duraciones similares han sido reportadas para aproximadamente las mismas condiciones en algodón (6) y frijol (14), incluyendo pequeñas variaciones entre huevos puestos sobre diferentes especies de plantas hospederas (25). La fase de huevo es quizás la menos afectada por la planta hospedera, ya que durante ella, el insecto no se alimenta de la planta por lo cual resulta difícil acertar las razones para estas diferencias de duración. No obstante, no deben descartarse efectos de microclima consecuencia de la arquitectura de cada especie de hospedera. Esto sugiere que el sustrato (especie de planta x edad de hoja) podría afectar la duración del período de huevo.
Aparentemente, la temperatura a la cual el insecto es criado, es el factor determinante de las mayores variaciones de duración de esta y otras fases del desarrollo del insecto (6, 25).
El cuadro 1 presenta las duraciones totales de la fase ninfal (N1-N4=eclosión del huevo- emergencia del adulto) expresadas en días, sobre las diferentes hospederas. No fueron detectadas diferencias estadísticamente significativas, pero el período de desarrollo ninfal mostró cierta variación de acuerdo a las especies de plantas. Aunque en promedio, el insecto tardó más en desarrollarse sobre tomate, allí hubo menor variación que en las otras hospederas (entre aproximadamente 4 y 5 veces menos). Coudriet et al. (12) a pesar que encontraron marcadas diferencias en la duración del desarrollo (huevo- emergencia del adulto; 17 % entre valores extremos) de B tabaci sobre varias especies de hospederas silvestres en el laboratorio, aquellas menos favorables fueron capaces de mantener apreciables poblaciones del insecto en el campo durante todo el año. Por lo tanto la duración del desarrollo puede no ser por si solo un factor determinante así como tampoco un buen indicador de la relación planta-artrópodo.
Cuadro 1. Duración de la fase ninfal de Bemisia tabaci, sobre hojas de cinco especies de plantas hospederas, en el laboratorio.
| Hospedera | Duración (días) Media±DS | # Unidades experimentales | # Individuo |
| Tomate | 15.5 ± 1.1a | 33 | 324 |
| Caraota | 15.4 ± 4.4a | 29 | 184 |
| Cayena | 14.7 ± 5.2a | 32 | 138 |
| Navidad | 14.6 ± 4.3a | 29 | 187 |
| Algodón | 13.9 ± 4.4a | 35 | 141 |
Comparaciones de medias realizadas mediante la prueba de Tukey. a: Medias seguidas de letras diferentes difieren significativamente (P < .05). DS= Desviación estandard.
El promedio general para la duración de la fase de ninfas, fue 14.8 ± 4.2 días, lo cual es menor que los obtenidos por otros autores (14, 25). Estas diferencias pueden deberse a las temperaturas ligeramente mayores bajo las cuales se realizó esta investigación (27.1 ± 1.3 versus 25.7 y 25 °C). No obstante, otros factores, tales como variaciones debido a razas o especies del insecto (3, 4), edad de las plantas hospederas (8), etc., no deben ser descartadas. La duración de las ninfas en tomate y algodón tendió a ser uniforme entre las diferentes edades de hojas, en caraota hubo menor duración en las hojas maduras, mientras que para cayena y navidad la duración fue menor en las hojas senescentes. Aunque la duración total de la fase de ninfa (N1N4), no mostró diferencias estadísticamente significativas entre hospederas ni edades de hojas, al discriminar por estadíos, fueron detectadas diferencias para la duración de N3 entre las hojas jóvenes (3.2 ± 0.5) y las senescentes (2.8 ± 0.3) (P < .05).
Longevidad de los adultos. El promedio general de longevidad de los adultos, incluyendo ambos sexos, fue de 9.2 ± 3.0 días. No se encontraron diferencias estadísticamente significativas por efecto de las especies de plantas hospederas. El orden descendente de longevidad fue algodón, tomate, caraota, navidad y cayena (amplitud de variación: 9.8 ± 3.7 a 8.4 ± 3.2). En tomate, algodón y cayena, la longevidad de los adultos decreció con la edad de las hojas. En caraota, la mayor longevidad ocurrió sobre hojas maduras, siendo lo contrario en navidad.
El cuadro 2 muestra las duraciones de las fases y estadíos de desarrollo, así como longevidad de los adultos de la MBT, discriminadas por sexo. No hubo diferencias significativas entre sexos para las diferentes fases y estadíos a excepción de la longevidad de los adultos, donde las hembras duraron más que los machos (P < .01). Las longevidades de los adultos reportadas en la literatura, difieren de acuerdo a la temperatura, humedad relativa y hospedera sobre la cual se desarrollo y/o alimentó (6, 14, 25).
Supervivencia hasta adulto. En promedio general, el 69.23 ± 8.8 % de individuos sobrevivió desde huevo recién puesto hasta adulto, variando sin significancia estadística entre las especies de plantas hospederas (tomate > cayena > algodón > caraota > navidad; amplitud: 79.40 ± 18.6 y 56.06 ± 24.79 %). La supervivencia puede variar de acuerdo a las especies de plantas hospederas evaluadas (8, 31) y la edad de la planta (8). Sobre hojas de tomate y caraota tendió a aumentar con la edad de la hoja; para cayena y navidad, en las hojas maduras fue menor que en las jóvenes y senescentes, siendo lo contrario en algodón. La diferncia entre hojas senescentes y jóvenes en tomate resultó significativa (P < .05). En algunos estudios de biología de la MBT no se hace referencia a la supervivencia del insecto (6, 12, 14, 25); sin embargo este es un factor importante, ya que la capacidad de un artrópodo para desarrollar poblaciones sobre una determinada planta hospedera, depende en parte de cuantos sobreviven para reproducirse sobre ella.
Cuadro 2. Duración promedio de los estadíos de Bemisia tabaci, discriminados por sexo, sobre hojas de tres edades y cinco especies de plantas hospederas, en el laboratorio.
| Variables | Sexo | Duración Media ± DS | GL | Prob > t |
| Huevo | Hembra | 6.5 ± 0.9 | 260 | 0.57ns |
| Macho | 6.5 ± 0.8 | |||
| N1 | Hembra | 3.8 ± 0.6 | 258 | 0.80ns |
| Macho | 3.8 ± 0.7 | |||
| N2 | Hembra | 4.0 ± 0.8 | 252 | 0.43ns |
| Macho | 4.1 ± 0.8 | |||
| N3 | Hembra | 3.2 ± 0.6 | 250 | 0.71ns |
| Macho | 3.2 ± 0.6 | |||
| N4 | Hembra | 2.7 ± 0.5 | 249 | 0.91ns |
| Macho | 2.7 ± 0.5 | |||
| Pupa | Hembra | 2.1 ± 0.3 | 235 | 0.85ns |
| Macho | 2.1 ± 0.3 | |||
| Adulto | Hembra | 10.9 ± 2.6 | 221 | 0.0001** |
| Macho | 8.3 ± 1.9 |
Comparaciones de medias realizadas mediante la prueba de Student (P < .01). GL = Grados de libertad.
Proporción de sexos. En general, la proporción de sexos fue de 1.58 ± 0.22: 1 (hembra:macho), sin mayores variaciones entre plantas hospederas, lo cual se asemeja a los reportados por otros autores (14, 25). Al ser comparadas las edades de hojas, sin discriminar entre hospederas, se observa una mayor proporción de hembras producidas en las hojas jóvenes (1.81 ± 0.43:1) con respecto a las encontradas en las hojas maduras (1.5 ± 0.34:1) y senescentes (1.44 ± 0.22:1). No obstante esa tendencia varió con las especies de plantas, siendo tomate y caraota relativamente uniforme con proporciones de hembras ligeramente mayores en las hojas maduras. De todos modos, el efecto de esa diferencia (20.67 y 25.69 % más de hembras), podría tener significancia en el desarrollo de las poblaciones del insecto cuando la planta se encuentra en pleno desarrollo vegetativo, debido a que favorecería la tasa de natalidad y consecuentemente el aumento poblacional durante esa fase de la fenología de la planta.
Fertilidad de las hembras. La fertilidad fue en promedio general de 42.2 ± 19.6 huevos/hembra, detectándose diferencias estadísticamente significativas (P < .05) entre las plantas de tomate, caraota y navidad, con respecto a cayena y algodón, con 1.9 veces más huevos en tomate que en algodón (cuadro 3). Estos resultados, corroboran la preferencia que muestran los adultos por oviponer en las plantas de hojas pubescentes(7, 14, 25), ya que la cayena y el algodon aquí utilizados, son de superficie foliar muy lisa al contrario de las otras tres especies.
Al comparar edades de hojas sin discriminar por hospederas, se observó mayor fertilidad en hembras provenientes de/y oviponiendo sobre hojas maduras (46.3 ± 10.0), seguida de las senescentes (41.3 ± 16.1) y jóvenes (30.0 ± 8.6), encontrándose diferencias estadísticamente significativas (P < .05) entre las hojas maduras y jóvenes. En esta respuesta están confundidos los efectos de fertilidad de las hembras con su aceptación del sustrato para oviponer. No obstante, es evidente la mayor fertilidad de hembras provenientes de hojas maduras, ya que al iniciar las cohortes evaluadas en este experimento, las hembras confinadas sobre hojas jóvenes pusieron mayor número de huevos que aquellas sobre hojas maduras y senescentes (información a ser publicada aparte).
Tiempo generacional. En las condiciones de este estudio, sin discriminar las especies de plantas hospederas, una generación de MBT (huevohuevo) puede completarse en 26.2 ± 7.2 días. El orden decreciente fue: tomate (28.9 ± 0.8), cayena (26.5 ± 9.1), caraota (25.7 ± 7.6), navidad (25.7 ± 7.4) y algodón (25.4 ± 8.1), sin haber significancia estadística. En tomate hubo menos variabilidad, comparado con las otras hospederas (amplitud: 8.8 a 10.8 veces menos). Estos resultados sugieren que la MBT puede tener entre 13 y 14 generaciones/año sobre las especies de plantas estudiadas, aproximadamente de 4 a 5/ciclo de cultivo en hospederas de 120 días, lo cual entra en el marco de los resultados de otros estudios (20). Aunque T está muy relacionado con la duración del desarrollo, expresa mejor la relación temporal del insecto con la planta, puesto que incluye la longevidad del adulto y su secuencia reproductiva.
Cuadro 3. Fertilidad promedio de las hembras de Bemisia tabaci, criadas sobre hojas de cinco especies de plantas hospederas, en el laboratorio.
| Hospedera | # de huevos/hembra | # Unidaes | # Hembras |
| Media ± DS | experimentales | ||
| Tomate | 53.17 ± 10.01a | 33 | 187 |
| Caraota | 49.97 ± 17.72a | 26 | 110 |
| Navidad | 48.81 ± 22.52a | 29 | 112 |
| Cayena | 30.86 ± 14.25b | 31 | 83 |
| Algodón | 28.03 ± 15.91b | 34 | 50 |
Comparaciones de medias realizadas mediante la prueba de Tukey. a, b: Medias seguidas de letras diferentes no difieren significativamente (P < .05). DS = Desviación estandard.
En tomate y algodón el tiempo generacional no varió con la edad de la hoja. En cayena se observó una disminución en la medida que las hojas envejecieron. Mientras que en caraota y navidad se observó lo contrario.
Tasa neta reproductiva (Ro). En promedio general, al cabo de una generación, las poblaciones de la MBT podrían multiplicarse 17.9 ± 5.8 veces (individuos/individuo); siendo en tomate y caraota 2.12 y 1.6 veces mayores que en cayena y algodón (P < .05); navidad no difirió de ninguno de los dos grupos (cuadro 4). En consecuencia las poblaciones de MBT sobre este cultivar de tomate, podrian duplicar a aquellas sobre algodón en términos de una generación. La tendencia fue similar a la observada en la fertilidad de las hembras, lo cual sugiere que este sería el factor con mayor efecto en las diferencias población-planta hospedera observadas en el campo .
Tasa intrínseca de desarrollo poblacional (rm). En promedio general, un individuo de MBT puede aportar 0.095 ± 0.02 indivíduos por día a lo largo de toda su vida. El potencial de desarrollo poblacional resultó significativamente mayor (P < .05) sobre tomate, comparado con algodón y cayena; caraota y navidad no difirieron de ambos grupos (cuadro 5). La figura 1, muestra el resultado de la simulación del potencial de desarrollo poblacional durante los primeros 90 días del inicio de la infestación, basado en los valores de rm obtenidos para cada especie de planta hospedera. En ese lapso, sobre tomate las poblaciones podrían alcanzar niveles aproximadamente 4, 5, 21 y 27 veces mayores que sobre caraota, navidad, algodón y cayena, respectivamente.
El efecto de la edad de la hoja no mantiene la misma tendencia para todas las hospederas. En tomate y caraota, los valores de rm aumentan ligeramente con el envejecimiento de las hojas, en navidad y cayena ocurre lo contrario. En algodón, las hojas maduras parecen ser las más favorables para el insecto. Solo resulto significativa (P < .05) la diferencia entre hojas jóvenes y senescentes de navidad. Ello sugiere que dependiendo de la especie de planta, el desarrollo de poblaciones de la MBT puede variar de acuerdo a la fenología de la misma; en lo cual incide la edad de hoja predomi-nante en cada etapa de su ciclo. En lechuga, Lactuca sativa L. la oviposición y la supervivencia de ninfas han sido encontrado inversamente relacionado con la maduración de las hojas, a pesar que las ninfas alcanzaron el floema con su estilete bucal, lo cual sugiere que algún otro factor mas allá de la posibilidad física para alimentarse, influyó en la mortalidad (8).
Cuadro 4. Tasa neta reproductiva (Ro) de Bemisia tabaci, sobre hojas de cinco especies de plantas hospederas, en el laboratorio.
| Hospedera | Ro Media ± DS | # Unidades experimentales | # Individuos |
| Tomate | 25.9 ± 6.3a | 33 | 430 |
| Caraota | 20.6 ± 9.1a | 26 | 304 |
| Navidad | 18.2 ± 8.2ab | 28 | 302 |
| Cayena | 12.7 ± 6.4b | 30 | 193 |
| Algodón | 12.2 ± 6.6b | 34 | 218 |
Comparaciones de medias realizadas mediante la prueba de Tukey. a, b: Medias seguidas de letras diferentes no difieren significativamente (P < .05). DS = Desviación estandard.
Cuadro 5. Tasa intrínseca de desarrollo poblacional (rm) de Bemisia tabaci, sobre hojas de cinco especies de plantas hospederas, en el laboratorio.
| Hospedera | rm Media ± DS | # Unidades experimentales | # Individuo |
| Tomate | 0.116 ± 0.01a | 33 | 430 |
| Caraota | 0.102 ± 0.03ab | 26 | 304 |
| Navidad | 0.098 ± 0.03ab | 28 | 302 |
| Algodón | 0.082 ± 0.03b | 34 | 193 |
| Cayena | 0.079 ± 0.03b | 30 | 218 |
Comparaciones de medias realizadas mediante la prueba de Tukey. a, b: Medias seguidas de letras diferentes no difieren significativamente (P < .05). DS = Desviación estandard.
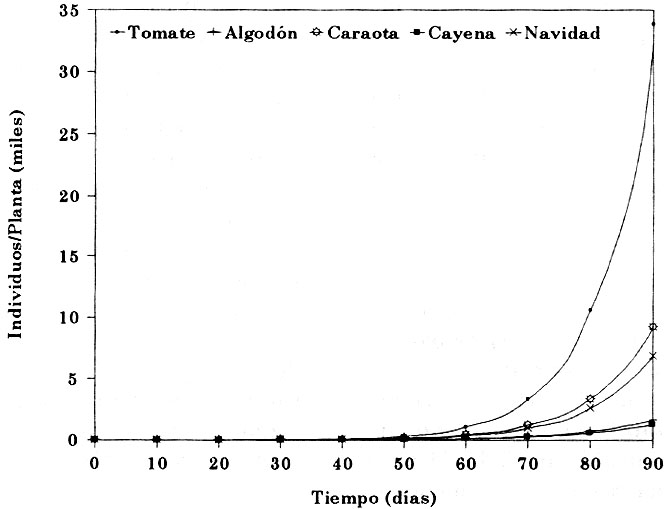
Figura 1. Simulación del desarrollo poblacional de Bemisia tabaci creciendo sobre cinco especies de plantas hospederas en condiciones de laboratorio.
En general, los resultados muestran consistencia como indicadores de cuan favorables son las diferentes especies de plantas hospederas para la raza de B. tabaci, evaluada durante este experimento. No obstante, cada una de las variables considerada por separado, no siempre deja clara evidencia de ello. Los márgenes de variabilidad observados, con frecuencia enmascaran los efectos de la hospedera sobre el insecto. Los cambios en la calidad de las plantas dentro del ambiente de cría, varió de acuerdo a cada especie. Esto, junto con posibles cambios en la colonia debido a la presión de selección por las condiciones de cría, pudo haber aumentado la variabilidad en el tiempo.
La duración del desarrollo no siempre resulta buen indicador de lo favorable que las condiciones de cría resultan para el artrópodo. En este caso, la aceleración del desarrollo debido a la planta, ha sido acompañado por menor supervivencia y/o fertilidad dependiendo del caso.
La supervivencia refleja cierta diferencia, sin embargo, la variabilidad dentro de especies de plantas, no permite precisarlas estadísticamente. Aunque de acuerdo a los promedios de algunas de las variables medidas (duración del desarrollo y tiempo generacional) tomate se muestra menos favorable para la MBT, la variabilidad resultó mucho menor que sobre las otras plantas hospederas, lo cual puede interpretarse como mejor adaptación a esa especie de planta. La fertilidad, denota diferencias acordes con el orden de favorabilidad de las especies de plantas (tomate, caraota, navidad > cayena y algodón). Finalmente, el efecto global de los resultados parciales de las variables evaluadas son adecuadamente resumidos por Ro y rm, las cuales concretan las diferencias de relación insecto-planta, cuyas proyecciones poblacionales pueden apreciarse con las simulaciones en base a rm (figura 1).
Estos resultados encajan con ciertos resultados de investigaciones de campo. Las altas infestaciones por MBT entre otros artrópodos, que se desarrollan en tomate bajo uso continuado de insecticidas (10, 17) evidencian que cuando algún factor interfiere con la acción de los enemigos naturales, los rápidos aumentos poblacionales del fitófago muestran lo adecuado que la planta resulta para el insecto, además de cualquier otro factor. Sería conveniente desarrollar algún modelo estadístico para comparar las proyecciones de rm al cabo del tiempo.
Literatura citada
1. Andrewartha, H.G. y L.C., Birch.1954. The Distribution and Abundance of Animals. University of Chicago Press, Chicago.
2. Arnal, E., L. Russel y E. Debrot. 1993. Plantas hospederas de la mosca blanca Bemisia tabaci (Gennadius) en Venezue. Agron. Trop. (Maracay) 43(5-6):267-285.
3. Barinaga, M. 1993. Is devastating whitefly invader really a new species? Science 259: 30.
4. Bellow, T.S., T.M. Perring, R.J. Gill y D.H. Hedrick. 1994. Description of a species of Bemisia (Homoptera: Aleyrodidae). Ann. Entomol. Soc. Am. 87:195-206).
5. Bentz, J., J. Reeves, P. Barbosa y B. Francis. 1995. Effect of nitrogen fertilizer source and level on ovipositional choice of poinsettia by Bemisia argentifolii (Homoptera: Aleyrodidae). J. Econ. Entomol. 88(5): 1388-1392.
6. Butler, G.D., jr., T.J. Henneberry y T.E. Clayton. 1983. Bemisia tabaci (Homoptera: Aleyrodidae): development, oviposition, and longevity in relation to temperature. Ann. Entomol. Soc. Am. 76(2):310-313.
7. Butler, G.D., jr., T.J. Henneberry y F.D. Wilson. 1986. Bemisia tabaci (Ho-moptera: Aleyrodidae) on cotton: adult activity and cultivar oviposition preference. J. Econ. Entomol. 79(2):350-354.
8. Byrne, D. N. 1989. Effect of plant maturity on oviposition and nymphal mortality of Bemisia tabaci (Homoptera: Aleyrodidae). Environ. Entomol. 18(3): 429-432.
9. Chi, H. y H., Liu. 1985. Two new methods for the study of insect population ecology. Bull. Inst. Zool. Acad. Sinica. 24: 225-240.
10. Chirinos, D. T., F. Geraud-Pouey. 1996. Efectos de algunos insecticidas sobre la entomofauna del tomate en el noroeste del estado Zulia, Venezuela. Interciencia 21(1): 31-36.
11. Clark, L.R., P.W., Geier, R.D., Hughes y R.F., Morris. 1967. The Ecology of Insect Populations in Theory and Practice. Metheun & Coltd, Londres.
12. Coudriet, D.L., D.E. Meyerdirk, N. Prabhaker y A.N. Kishaba. 1986. Bionomics of sweetpotato whitefly (Homoptera: Aleyrodidae) on weed hosts in the Imperial Valley, California. Environ. Entomol. 15(6): 1179-1183.
13. Debrot, E., F. Herold y F. Dao. 1963. Notas preliminas sobre el mosaíco amarillento del tomate en Venezuela. Agron. Trop. (Maracay) 13(1): 33-41.
14. Eichelkraut, K. y C. Cardona. 1989. Biología, cria masal y aspectos de la Mosca Blanca Bemisia tabaci (Gennadius) (Homoptera: Aleyrodidae), como plaga del Frijol Común. Turrialba 39(1): 55-62.
15. Falcon, L.A. 1971. Progreso del control integrado en el algodón de Nicaragua. Rev. Peru. Entomol. Agr. 14(2): 376-378.
16. Geraud-Pouey, F., D. Chirinos y G.Rivero. 1995. Artrópodos asociados con el cultivo del tomate en Venezuela. Boletín Entomol. Venez. N.S. 10(1):31-49
17. Geraud-Pouey, F.; D. T. Chirinos y J. A. Vergara. 1996. Efectos colaterales de tratamientos con insecticidas sobre la entomofauna del tomate, Lycopersicon esculentum Miller, cv. Peto Seed 98, en la zona del Río Limón, Estado Zulia, Venezuela. Rev. Fac. Agron. (LUZ). 13(3): 313-325.
18. Greathead, A. 1986. Host plants. Cap. 3 en M.J.W. Cock (ed.). Bemisia tabaci a literature survey on the cotton whitefly with an annotated bibliography. FAO, CAB International.
19. Guagliumi, P. 1966. Insetti e Arcnidi delle Piante Comuni del Venezuela Segnalati nel Periodo 1938-1963. Instituto Agronomico per L'Otramare, Florencia.
20. Husain, M.A. y K.N. Trehan. 1933. Observations on the life-history, bionomics and control of the whitefly of cotton (Bemisia gossypiperda M.& L.). Indian J. of Agric. Sci. 3: 701-753.
21. King, B. y J. Saunders. 1984. Las plagas invertebradas de cultivos anuales alimenticios en América Central. Overseas Development Administration, Londres.
22. Krebs, C. 1981. Ecology: The experimental analysis of distribution and abundance. 2nd. edit. Harper and Row. New Yord.
23. Lastra, J. y R. de Uscátegui. 1975. Viruses affecting tomatoes in Venezuela. Phytopathologische Zeitschrift 84: 253-258.
24. Liu, T. y P.A. Stansly. 1995. Oviposition by Bemisia argentifolii (Homoptera-Aleyrodidae) on tomato: effects of leaf factors and insecticides residues. J. Econ. Entomol. 88(4): 992-997.
25. López-Avila, A., 1986. Taxonomy and biology. Cap. 1. en M.J.W. Cock (ed.). Bemisia tabaci a Literature Survey on the Cotton Whitefly with an Annotated Bibliography. FAO, CAB International.
26. Price, P., C. Bouton, P. Gross, B. McPheron, J. Thompson y A. Weis. 1980. Interactions among three trophic levels: influence of plants on interactions between insect herbivores and natural enemies. Annu. Rev. Ecol. Syst. 11:41-64.
27. SAS Institute Inc. 1989. Paquete Estadístico para microcomputadoras. Versión 6. Cary, N.C.
28. Simmons, A. M. 1994. Oviposition on vegetables by Bemisia tabaci (Homoptera: Aleyrodidae): Temporal and leaf surface factors. Environ. Entomol. 23(2): 381-389.
29. Steel, R. G. y J. H. Torrie. 1960. Principles and Procedures of Statistics. McGraw-Hill, New York.
30. Uzcátegui, R. de y R. Lastra. 1978. Transmission and physical properties of the causal agent of mosaico amarillo del tomate (tomato yellow mosaic). Phytopathology 68:985-988.
31. Zalom, F. G., C. Castañé y R. Gabarra. 1995. Selecction of some winterspring vegetable crop hosts by Bemisia argentifolii (Homoptera: Aleyrodidae). J. Econ. Entomol. 88(1): 70-76.